
CDBからのニュース、お知らせを掲載しています。
インスリン/インスリン様成長因子(IGF)シグナルは成長・代謝・生殖・加齢などに重要で、進化的に広く保存されていることが知られる。哺乳類では、主に代謝に働くインスリンと体の成長に関わるIGFが各役割を分担するが、ショウジョウバエでは8つのDilp(Dilp1~8)がこれらの働きを担う。これら内分泌ホルモンの機能は環境の変化、特に個体が取り込む栄養の状態に応じて柔軟に調節される必要があるが、その分子メカニズムについては未だ謎が多い。
理研CDBの岡本直樹元研究員(成長シグナル研究チーム、西村隆史チームリーダー)らは、ショウジョウバエをモデルに、成長制御ホルモンdilp5が栄養状態に依存して発現制御される分子機構を明らかにした。この成果は、科学誌Developmental Cellに11月9日付で掲載された。なお、岡本元研究員は現在、カリフォルニア大学リバーサイド校に籍を移して研究を行なっている。
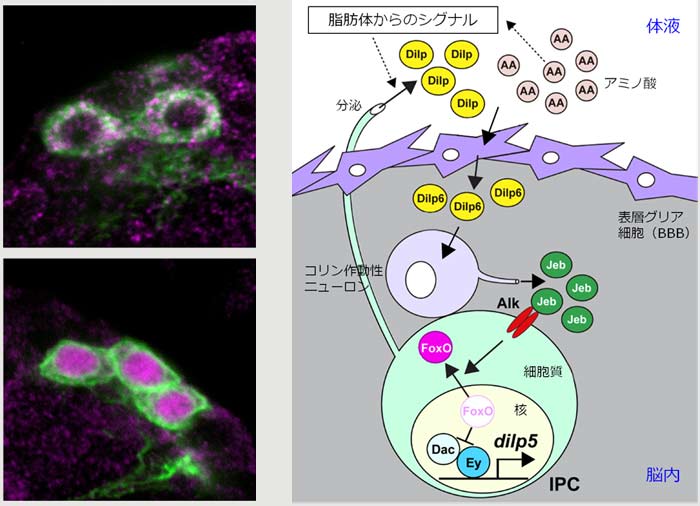
昆虫のDilpは、摂食に応じて放出される脂肪体(哺乳類の肝臓にあたる組織)からのシグナルを受けて、主に脳にあるインスリン産生細胞(IPC)から体液中に分泌されて全身を巡る。中でもDilp5はIPCで産生される主要なDilpの1つで全身の成長に重要な働きを示す。dilp5は栄養状態に応じて発現が大きく変動することが知られており、実際に孵化直後の摂食開始に反応するように幼虫初期から発現が急激に上昇する。岡本らはこれまでに、dilp5の発現が2つの転写因子Dachshund(Dac)とEyeless(Ey)の協調作用によって活性化されることを明らかにしていた(*科学ニュース2012.2.10)が、dilp5の発現変動と栄養状態との間をつなぐ分子機構は不明だった。
岡本らはまず、dilip5の発現制御に作用する栄養素がアミノ酸であることを確認。アミノ酸センサーとしてよく知られるTorシグナルを、IPCにおける上流因子の第一候補として解析してみたところ、Torシグナル関連分子を発現低下させてもdilp5に影響が見られないことが分かった。dilp5の発現制御機構は、そう単純なものではないらしい。そこで、DacとEyを含む複合体の構成因子の中で栄養状態によって変化するものを探索した。その結果、飢餓状態になると、IPCにおいて転写因子FoxOが細胞質から核内に移行することが分かった。さらに、核内移行したFoxOはEyと直接結合してDacとEyの結合と拮抗することが示され、FoxOの細胞内局在がdilp5の発現制御のON/OFFを切り替えることが明らかになった。
では、FoxOの局在はどのように制御されるのだろうか。種々のシグナル経路関連分子を探索した結果、Alk(anaplastic lymphoma kinase:受容体型チロシンキナーゼ)の活性化がFoxOを細胞質に局在させ、dilp5の発現を促進することが分かった。このAlkを活性化するのは分泌型リガンドJelly belly(Jeb)であり、Jebが産生されるのはコリン作動性ニューロンだった。IPCに近接するコリン作動性ニューロンがJebを産生・放出することで、遠隔操作的にFoxOの局在が制御されるのだ。さらにJebの上流因子を辿ると、Jebの機能は中枢神経組織の表層に位置するグリア細胞から産生されるDilp6によって制御されていることが判明した。昆虫の表層グリア細胞は、哺乳類の血液脳関門と同様の働きを担っており、体液とも直接触れ合う。そこでさらなる上流因子を探索した結果、dilp6の発現は体液中に放出されたIPC由来の他のDilp(Dilp2, 3, 5)によって誘導されることを突き止めた。
摂食に応じてIPCから産生・放出されたDilpが、複数の細胞種が産生する種々の分子のシグナルリレーとポジティブフィードバック機構を経て増幅される。本研究から、dilp5の発現を栄養依存的にダイナミックに変化させる、複雑な分子機構の全体像が解明された。「それにしても、ショウジョウバエはなぜこんなにもややこしいメカニズムを採用しているのか。私たちはdilp5の変異体を詳細に解析し、低栄養状態下における成長率および成体の体サイズが野生型と比べて減少し、さなぎに至るまでの発育時間も長くなることを見出しました。dilp5の複雑な発現制御機構は、低栄養状態でもインスリンシグナルを維持して体の成長を推し進めるために重要なのだと考えられます」と西村チームリーダーは語る。「転写因子FoxOの機能やコリン作動性ニューロンの寄与は哺乳類インスリンの制御にも共通することから、今回明らかにした分子機構は進化的に広く保存されている可能性があります。環境変化に順応する生物たちの巧みな生存戦略に今後も迫っていきたい。」
| 掲載された論文 | |
|---|---|
| 関連記事 |
![CDB 理化学研究所 発生・再生科学総合研究センター [RIKEN CENTER FOR DEVELOPMENTAL BIOLOGY]](http://www.cdb.riken.jp/wp-content/themes/cdb/images/common/fLogo2.png)
〒650-0047 兵庫県神戸市中央区港島南町2-2-3
TEL : 078-306-0111
FAX : 078-306-0101
E-mail : cdb[at]cdb.riken.jp [at]を@に変えてメールしてください。

