RIKEN Center for Developmental Biology
2003 Annual Report
|
|
RIKEN Center for Developmental Biology 2003 Annual Report |
Laboratory for Vertebrate Axis Formation
|
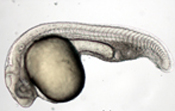
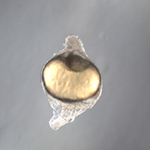
Vertebrate embryogenesis begins with a seemingly formless egg that, through processes of division, growth, differentiation, migration and rearrangement, rapidly gives rise to a highly organized structure characterized by a number of definitive axes. This transformation is particularly striking and rapid in the zebrafish (Danio rerio), which in the space of a single day develops from an ovum to a recognizably vertebrate body laid down in respect to multiple polar vectors. The speed of its growth and reproduction, coupled with its amenability to genetic studies and the revealing pellucidity of its embryo, make this organism an ideal system for the study of axis formation in early vertebrate development.
Masahiko Hibi's laboratory focuses on the molecular genetic cascade of events stemming from organizing centers in the zebrafish embryo. These centers, which emerge soon after fertilization, play pivotal roles in setting up positional axes; Hibi's lab is particularly interested in the formation of the dorsal-ventral (back-belly) axis, an essential step in the establishment of the body plan. An embryonic region known in zebrafish as the dorsal organizer guides dorsoventral patterning by generating inductive signals, including inhibitors of BMPs and Wnts (molecules which ultimately determine dorsal and ventral fates). These molecules play roles in a complex network of axis-determining factors, and the interplay between dorsalizing and ventralizing factors is central to the formation of the dorsal mesoderm and endoderm, and the neuroectoderm.
Hibi and colleagues set out to clone the ogon gene (which is also known as mercedes and short tail), in order to determine its precise relationship with Chordin, and the molecular bases for its action in regulating the establishment of the dorsal-ventral axis. The results of that positional cloning showed that ogon encodes a homolog of the Secreted Frizzled (Sizzled) protein, which has similarities to the Wnt receptor, Frizzled. The canonical Wnt pathway is known to play a role in axis formation, but it has been suggested that Ogon/Sizzled influences the dorsal-ventral axis independently of its effect on Wnt. To test this idea, members of the Hibi lab compared the activity of Ogon/Sizzled with that of Wnt inhibitors in ectopic expression studies, and found that the effects of Ogon/Sizzled misexpression resembled those of BMP, but not Wnt, inhibitors, suggesting an exceptional mode of action.
They next looked at the possible involvement of Chordin (which is known to be essential in the process of dorsalization) in Ogo/Szl’s effects by injecting ogon/sizzled RNA into mutants (chordino) lacking a functional chordin gene. This failed to rescue the ventralized phenotype of homozygous chordino mutants, indicating that Ogon/Sizzled activity depends on Chordin. The detailed functional relationship between these two dorsalizing factors remains to be worked out, but these initial findings suggest that Ogon/Sizzled augments Chordin activity either by inhibiting a Chordin inhibitor, by upregulating Chordin activity, or by making BMP more sensitive to Chordin’s inhibitory effects.
Neurogenesis Vertebrate neuronal tissues are generated in a stepwise fashion. These steps include neural induction, antero-posterior patterning, and neurogenesis. In amphibian and teleost (bony fish) embryos, neuroectoderm is induced by BMP inhibitors derived from organizing centers. Induced neuroectoderm is by default anterior in character, but a subset in some tissues is subsequently subject to posteriorizing transformations. The hindbrain and spinal cord, which are posteriorized regions, are specified by a signal from a region known as non-axial mesendoderm, while anterior neuroectoderm that does not receive this posteriorizing signal develops into fore- and midbrain. After these initial processes of neural induction and patterning, neurogenic regions, the domains in which neurogenesis takes place, are established.
The Hibi lab has been working to identify genes involved in the control of this patterning and neurogenesis, focusing on two genes in particular: the zinc finger gene fez-like expressed in the forebrain, and the posterior neuron-specific homeobox gene pnx. In collaborative work, they showed that fez-like is required for the formation of hypothalamic monoaminergic neurons in zebrafish. These neurons produce neurotransmitters such as dopamine and serotonin, which are centrally important to the regulation of mood, behavior, endocrine and cognitive functions in humans.
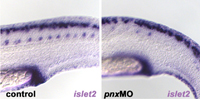
A separate study identified
a homeobox gene, pnx, involved
in the development of posterior neurons. This gene, which is regulated
by a signal from the posteriorizing |
|
Selected Publications
Levkowitz G, Zeller J, Sirotkin H I, French D, Schilbach S, Hashimoto H, Hibi M, Talbot W S and Rosenthal A. Zinc finger protein too few controls the development of monoaminergic neurons. Nat Neurosci 6:28-33 (2003).
Bae Y K, Shimizu T, Yabe T, Kim C H, Hirata T, Nojima H, Muraoka O, Hirano T and Hibi M. A homeobox gene, pnx, is involved in the formation of posterior neurons in zebrafish. Development 130:1853-65 (2003).
Yabe T, Shimizu T, Muraoka O, Bae Y K, Hirata T, Nojima H, Kawakami A, Hirano T and Hibi M. Ogon/Secreted Frizzled functions as a negative feedback regulator of Bmp signaling. Development 130:2705-16 (2003).
Shimizu T, Yamanaka Y, Nojima H, Yabe T, Hibi M and Hirano T. A novel repressor-type homeobox gene, ved, is involved in dharma/bozozok-mediated dorsal organizer formation in zebrafish. Mech Dev 118:125-38 (2002).